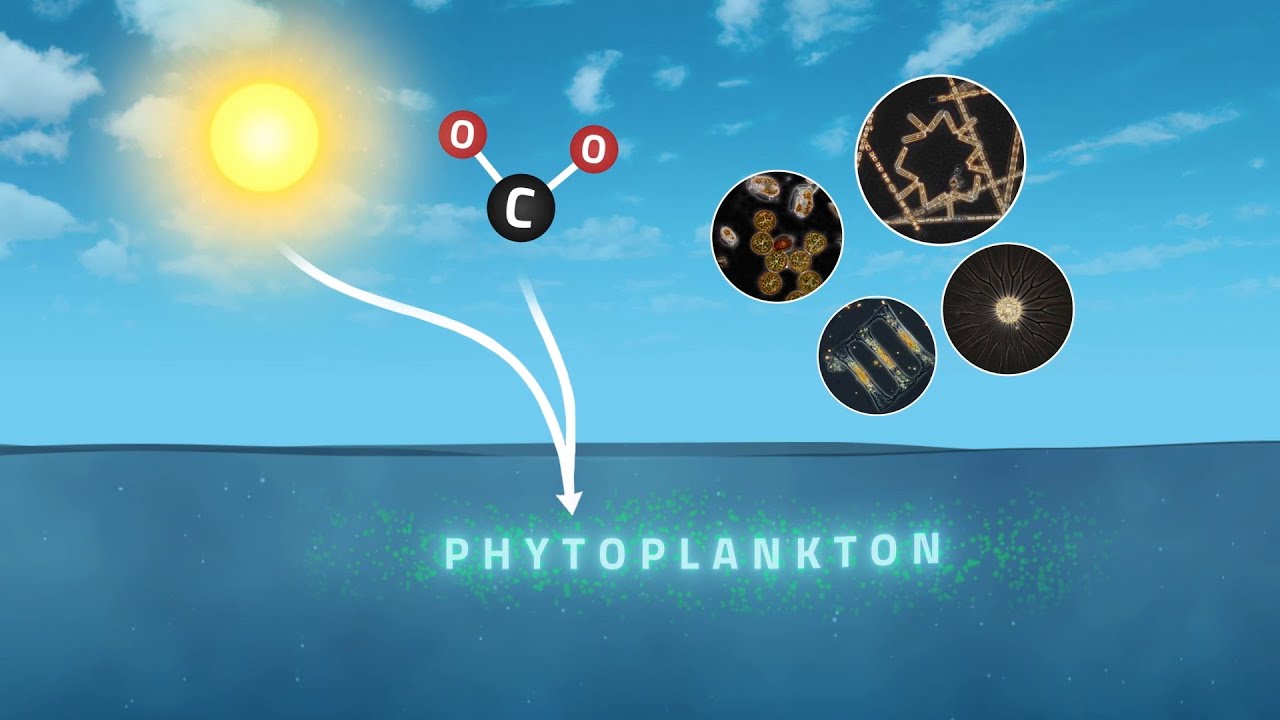
How Does the Biological Carbon Pump Work?
The biological carbon pump is the ocean's mechanism for transferring carbon from the atmosphere to the deep sea. It begins with photosynthesis: phytoplankton at the surface fix CO2 into organic matter. When they die, they aggregate into Marine Snow, a continuous shower of particles sinking from the sunlit zone to the abyss.
This is not a gentle drift. Marine Snow carries billions of tonnes of carbon below 1,000 meters annually, where it is locked away for centuries to millennia. Without this pump, atmospheric CO2 would be twice current levels.
Why Is the Microbial Loop the Hidden Engine?
Azam and colleagues discovered that bacteria process 50%% of all marine primary production through the Microbial Loop. Dissolved organic matter released by living and dying phytoplankton is consumed by bacteria, which are eaten by protists, which recycle nutrients back into the food web.
This loop is invisible but fundamental. It determines whether carbon stays at the surface (where it can re-enter the atmosphere) or gets packaged into particles dense enough to sink. The microbial loop is the decision point of the entire carbon pump.
What Is the Viral Shunt and Why Does It Matter?
Every liter of seawater contains 10 billion viruses. They kill 20-40%% of marine bacteria every single day, bursting cells and releasing their contents back into the dissolved organic matter pool. This viral shunt redirects carbon from higher trophic levels back into the microbial loop.
Far from being destructive, the viral shunt maintains microbial diversity (no species can dominate), recycles nutrients to the surface, and drives the evolution of microbial resistance. Marine viruses are ecosystem engineers, not pathogens.
Why Are Diatoms and Prochlorococcus So Different?
Diatoms are large (2-200 micrometers), glass-shelled, and dominant in nutrient-rich coastal waters. They sink fast and drive efficient carbon export. Prochlorococcus is tiny (0.5-1 micrometer), the most abundant photosynthesizer on Earth, and dominant in the nutrient-poor open ocean. It sinks slowly.
Together they produce approximately 40%% of Earth's oxygen. But they contribute to the carbon pump through fundamentally different mechanisms. The balance between diatom-driven export and cyanobacteria-driven recycling determines the ocean's carbon sequestration efficiency.
What Happens in the Twilight Zone?
The ocean twilight zone (200-1000 meters) is where the biological pump's fate is decided. BGC-Argo autonomous floats (Nature Geoscience, 2021) revealed that this zone processes far more carbon than previously estimated. Zooplankton migration, microbial respiration, and particle fragmentation all occur here.
Most Marine Snow is consumed or dissolved before reaching the deep ocean floor. Only 1-2%% of surface production survives to 1,000 meters. But that fraction is enormous at planetary scale, and it is this fraction that determines long-term climate.
How Are Microplastics Disrupting the Pump?
Microplastics interfere with Marine Snow formation by altering particle aggregation and sinking rates. Research suggests this could reduce the biological pump's efficiency by 10-25%%. Additionally, microplastics provide surfaces for bacterial colonization that redirect carbon processing away from sinking and toward surface recycling.
The plastics we dump on land end up disrupting the deepest carbon cycle on the planet.
How Much CO2 Has the Ocean Already Absorbed?
Since industrialization, the ocean has absorbed approximately 525 billion tonnes of anthropogenic CO2, roughly 30%% of all human emissions. This massive buffering effect has slowed climate change. But the cost is ocean acidification: as CO2 dissolves, it forms carbonic acid, lowering pH and threatening the calcifying organisms (corals, coccolithophores, pteropods) that are integral to the biological pump.
What Did BGC-Argo and Tara Oceans Discover?
The Tara Oceans expedition collected 35,000 samples across all ocean basins, discovering millions of unknown microbial genes and revealing that plankton community networks, not just abundance, predict carbon export efficiency (Guidi et al., Nature 2016).
BGC-Argo floats are now providing continuous, autonomous measurements of ocean biogeochemistry at depth. Together, these programs are rewriting our understanding of how the ocean breathes.
What Is the Mineral Ballast Hypothesis?
Not all Marine Snow sinks at the same speed. Armstrong et al. (2002) proved that mineral ballast — calcium carbonate from coccolithophore shells and silica from diatom frustules — increases sinking speed 10-100x. Particulate organic carbon flux correlates with mineral flux with R-squared of 0.8.
Without these heavy mineral shells, most organic carbon would be consumed by bacteria before reaching the deep ocean. Diatoms and coccolithophores are not just oxygen producers — they are the gravitational engines of the biological pump.
What Is Recalcitrant Dissolved Organic Carbon?
Jiao et al. (2010) in Nature Reviews Microbiology defined the Microbial Carbon Pump (MCP) — a process distinct from the biological pump. Microbes transform labile dissolved organic matter into Refractory DOC (RDOC) that persists for 4,000-6,000 years.
The RDOC reservoir holds 662 petagramsof carbon — the largest pool of reduced carbon on Earth. This 'invisible' carbon is chemically 'unreadable' to most organisms. The MCP may exceed the biological pump's sequestration in the vast oligotrophic ocean gyres.
Why Does Mixotrophy Change Everything?
Mitra et al. (2014) established that 50%% of marine protists are mixotrophic — combining photosynthesis and phagotrophy. They photosynthesize when light is available and eat bacteria when it is not. This dual metabolism dominates oligotrophic gyres.
Mixotrophy fundamentally changes carbon cycling models. Traditional models assume separate 'producers' and 'consumers.' In reality, the same organism does both, creating a metabolic flexibility that stabilizes the surface ocean food web and buffers it against nutrient limitation.
What Would Happen If the Biological Pump Failed?
Without the biological pump, atmospheric CO2 would be approximately twice current levels — returning the climate to a state not seen in 50 million years. The pump fails when surface warming increases stratification, trapping nutrients below the thermocline and starving phytoplankton.
Behrenfeld et al. (2006) in Nature showed this is already happening: ocean primary productivity has declined since 1999. The plankton that produce every second breath are being starved by the very warming they help buffer against. This feedback loop is the most consequential planetary risk that most people have never heard of.
What Connects the Ocean to Every Other System?
Rivers carry nutrients from the soil that fuel coastal blooms. Those blooms produce DMS that seeds clouds in the atmosphere. Ocean oxygen feeds the holobiont with every breath. The ethology of marine cooperation — cleaner fish markets, whale nutrient pumps — is the biological market theory of the deep.
What Happens in the Twilight Zone (200-1000m)?
The mesopelagic twilight zone is the most critical filter in the global carbon cycle. Every night, trillions of zooplankton migrate to the surface to feed and return to depth at dawn — the diel vertical migration, the largest movement of biomass on Earth. By feeding at the surface and excreting at depth, zooplankton inject carbon directly into the deep ocean, bypassing slow gravitational sinking.
Salps create the ocean's high-speed rail. Their membrane-bound fecal pellets sink at over 1,000 meters per day — so fast that bacteria cannot consume them in transit. This fast export is a critical climate buffer that ensures carbon reaches the seafloor for millennial storage.
How Does the Martin Curve Define Pump Efficiency?
The Martin Curve describes how organic carbon flux decreases with depth: F(z) = F(euphotic) x (z/z_euphotic) to the power of -b, where b is approximately 0.86 globally. This means 90%% of surface carbon is consumed by bacteria before reaching 1,000 meters. Only about 1%% reaches the seafloor.
Even a small shift in the b-value has massive climate consequences. If warming increases b (more efficient bacterial recycling), less carbon reaches the deep and more CO2 stays in the atmosphere. The plankton that start the cascade determine whether the pump works or fails.
How Does Mineral Ballast Accelerate Carbon Sinking?
Not all Marine Snow sinks at the same speed. Armstrong et al. (2002) proved that mineral ballast from coccolithophore calcium carbonate shells and diatom silica frustules increases sinking speed 10-100x. Heavy minerals drag organic carbon down faster than bacteria can consume it.
Dust from the air microbiome — Saharan and Gobi desert particles — provides additional lithogenic ballast. In years with high dust deposition, the Martin Curve b-value drops significantly because carbon races past the hungry twilight zone bacteria. The biological pump is not just biology. It is biogeochemistry — minerals and microbes working together.
How Do Giant Larvaceans Act as High-Speed Carbon Elevators?
Robison et al. (2005) in Science discovered that giant larvaceans build mucus houses up to 1 meter across that filter 10-40%% of upper ocean carbon per day. When clogged, the larvacean discards the house. The abandoned structure sinks at 800-1,000 meters per day — 10-100 times faster than individual particle settling. Henschke et al. (2019) quantified 100 million tons of carbon transported annually by these organisms.
What Is the Carbonate Counter-Pump Paradox?
Coccolithophores build calcium carbonate shells. The chemical reaction releases CO2: Ca2+ + 2HCO3- produces CaCO3 + CO2 + H2O. For every mole of carbon buried as carbonate, one mole is released as CO2. Yet their shells are the most efficient ballast mineral, increasing sinking speed to 100-200 meters per day.
The CO2 released during calcification is a short-term cost. The increased transport efficiency is a long-term gain. Ocean acidification weakens coccolithophore calcification — thinner, more fragile shells that reduce ballast efficiency at the same time warming increases stratification. Models disagree on whether the net feedback increases or decreases sequestration.
How Do Prochlorococcus Ecotypes Divide the Ocean by Light?
Johnson et al. (2006) revealed that Prochlorococcus is a family of distinct ecotypes. High-Light adapted (HL) ecotypes dominate the surface. Low-Light adapted (LL) ecotypes occupy 100-200 meters. Genomes range 1.6-2.4 megabases — the smallest of any photosynthetic organism. HL ecotypes cannot survive below 50 meters. LL ecotypes are outcompeted at the surface. This vertical partitioning maximizes total water column productivity.
Whether climate change will shift ecotype distributions is uncertain. Warming may expand HL habitat while increased stratification reduces nutrient supply to LL depths. The plankton-oxygen link depends on these dynamics.
How Do Marine Viruses Redirect the Carbon Cycle?
Sullivan et al. (2017) defined the dual role of ocean viruses. The viral shunt bursts bacterial cells, releasing their carbon as dissolved organic matter that stays at the surface — feeding the microbial loop instead of sinking. The viral shuttle does the opposite: infected cell aggregates become heavy and sink, exporting carbon to depth.
Which pathway dominates determines whether carbon stays in the atmosphere or reaches the deep ocean. Viruses kill 20-40%% of marine bacteria daily, but this is not destruction. It prevents any single species from monopolizing nutrients. Viral lysis maintains the microbial diversity that keeps the biological pump functioning across all ocean provinces.