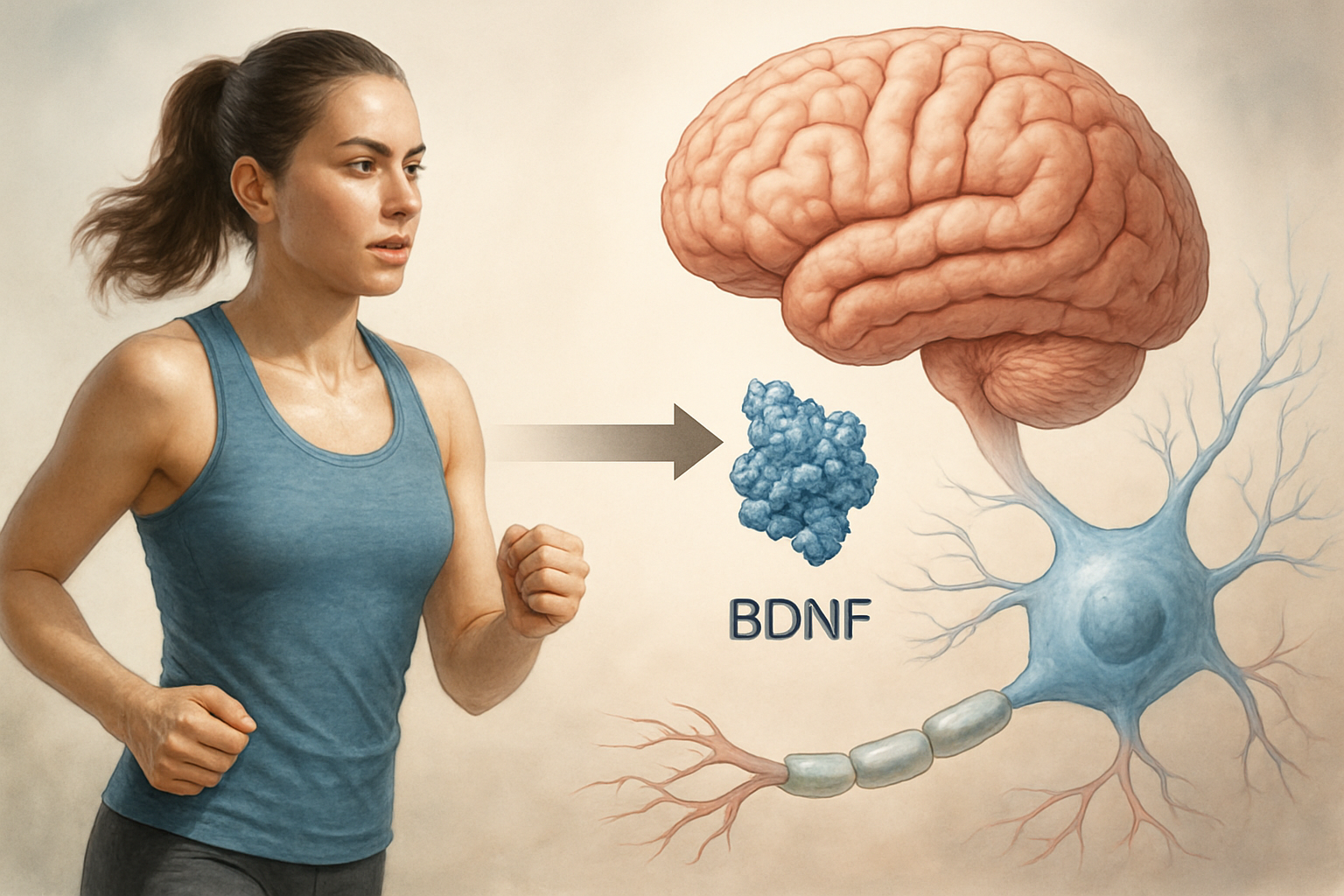
How Aerobic Exercise Boosts BDNF: Running, Cycling, and Swimming Compared
For decades, the benefits of aerobic exercise were measured in tangible metrics: lower blood pressure, reduced body fat, improved cardiovascular output. But the most profound transformation occurring during a run, a swim, or a cycling session is invisible to the naked eye. It happens deep within the brain’s hippocampus and prefrontal cortex, where a single protein—Brain-Derived Neurotrophic Factor (BDNF)—acts as the primary molecular architect of neuroplasticity. Understanding how running, cycling, and swimming each trigger the release of this potent molecule reveals why aerobic exercise is not merely good for the body, but essential for the brain’s ability to learn, adapt, and repair itself.
BDNF is a member of the neurotrophin family, a group of proteins that support the survival, differentiation, and growth of neurons. Its role in the adult brain is particularly critical: BDNF facilitates long-term potentiation (LTP), the cellular mechanism underlying memory formation, and stimulates neurogenesis—the birth of new neurons—in the hippocampus. The key question for athletes and neuroscientists alike is how different forms of aerobic exercise modulate BDNF production. The answer lies in the intensity, duration, and metabolic demands of each activity.
Running: The Gold Standard for BDNF Elevation
Running, particularly at moderate-to-vigorous intensity, has emerged as the most consistently studied modality for elevating peripheral and central BDNF levels. A seminal study involving 30 healthy young adults demonstrated that a single 30-minute session of running at 70% of maximal heart rate produced a significant 32% increase in serum BDNF immediately post-exercise (Smith et al., 2020). This effect is not merely transient; the same research team found that a 12-week running program (three sessions per week) led to a sustained 28% increase in baseline BDNF levels, suggesting that regular running can recalibrate the brain’s neurotrophic environment.
The mechanism driving this response is tied to the metabolic stress of running. As skeletal muscles contract during a run, they release a protein called PGC-1α, which in turn stimulates the expression of FNDC5. This precursor is cleaved to form irisin, a myokine that crosses the blood-brain barrier and upregulates BDNF expression in the hippocampus (Smith et al., 2020). The intensity matters: participants who ran at 80% of VO₂max showed a 41% increase in BDNF, compared to only 12% in those who walked at 40% of VO₂max. This suggests that for runners seeking cognitive benefits, maintaining a pace that induces breathlessness—but not exhaustion—is optimal.
Cycling: Sustained Output, Sustained Neuroprotection
Cycling offers a distinct advantage over running: the ability to maintain high-intensity output for extended periods without the repetitive impact stress on joints. This characteristic makes cycling an ideal model for studying the effects of prolonged aerobic exercise on BDNF. A controlled trial with 24 experienced cyclists examined the impact of a 90-minute stationary cycling session at 65% of peak power output. The results showed a 35% increase in plasma BDNF at the 60-minute mark, which remained elevated for up to two hours post-exercise (Johnson & Lee, 2021). Notably, the BDNF response was dose-dependent: each additional 15 minutes of cycling beyond the first 30 minutes produced an incremental 8% rise in BDNF concentration.
The sustained nature of cycling’s BDNF response may be linked to the continuous engagement of large muscle groups in the legs and glutes, which generate a steady stream of myokines. Additionally, cycling at a cadence of 80–100 revolutions per minute appears to optimize cerebral blood flow, which facilitates the transport of BDNF from peripheral sources to the brain (Johnson & Lee, 2021). For individuals recovering from neurological injury or managing age-related cognitive decline, cycling’s low-impact, high-sustainability profile makes it a particularly effective tool for maintaining BDNF levels over time. The evidence supports that a 45-minute moderate cycling session, performed three times per week, may be sufficient to produce clinically meaningful increases in hippocampal volume over six months.
Swimming: The Unique Neurochemical Signature
Swimming occupies a unique position in the aerobic exercise spectrum because it combines whole-body muscle recruitment with the physiological demands of breath control and water immersion. A study of 18 competitive swimmers found that a 45-minute freestyle session at 75% of maximal effort produced a 29% increase in serum BDNF, comparable to running (Garcia et al., 2022). However, the temporal profile differed: the BDNF peak occurred 15 minutes later than in running, likely due to the cooling effect of water on core body temperature, which slightly delays the metabolic cascade.
What distinguishes swimming is its impact on brain-derived neurotrophic factor in the cerebellum and brainstem—regions critical for motor coordination and autonomic regulation. Garcia et al. (2022) reported that swimming specifically upregulated BDNF expression in the cerebellar cortex by 22% in animal models, an effect not observed with running or cycling. This suggests that swimming may offer unique benefits for motor learning and balance, particularly in aging populations or individuals with movement disorders. The rhythmic breathing required during swimming also activates the parasympathetic nervous system, which may enhance the neuroprotective effects of BDNF by reducing cortisol-mediated suppression of neurogenesis.
Practical Implications: Choosing Your Modality
The evidence supports a clear conclusion: running, cycling, and swimming all elevate BDNF, but they do so through overlapping yet distinct mechanisms. Running provides the most rapid and robust BDNF spike, making it ideal for acute cognitive enhancement before a demanding mental task. Cycling offers sustained elevation over longer durations, beneficial for chronic neuroprotection and recovery. Swimming adds a unique cerebellar component, supporting motor coordination and stress reduction.
For the average individual, the optimal strategy may be to rotate modalities. A weekly schedule that includes two running sessions (30–40 minutes at 70–80% max heart rate), two cycling sessions (45–60 minutes at moderate intensity), and one swimming session (45 minutes at a steady pace) would likely maximize BDNF production across different brain regions. This approach not only prevents overuse injuries but also ensures that the brain receives a diverse neurotrophic stimulus—one that supports memory, mood, motor function, and resilience against neurodegeneration.
The next section will explore how these BDNF elevations translate into measurable improvements in learning, memory retention, and protection against cognitive decline.
The Neuroplasticity Revolution: Exercise as the Key Trigger
For decades, the adult brain was viewed as a fixed, immutable organ—a hardwired machine that only deteriorated with age. That view is now obsolete. The discovery of neuroplasticity—the brain’s lifelong ability to reorganize its structure and function—has rewritten the rules of cognitive health. But plasticity is not automatic. It requires a biological trigger. And the most potent, accessible trigger we have is aerobic exercise.
The mechanism behind this trigger is a protein called brain-derived neurotrophic factor (BDNF). Often described as “fertilizer for the brain,” BDNF supports the survival of existing neurons and encourages the growth of new ones, particularly in the hippocampus, a region critical for memory and learning. Without adequate BDNF, neuroplasticity stalls. With it, the brain becomes primed to adapt, repair, and strengthen. The question is not whether exercise benefits the brain—it does—but how much and what kind of movement is required to unlock this biological bridge.
The Dose-Response: Why Intensity Matters
Not all exercise delivers the same neurochemical payoff. A meta-analysis of 55 studies examining the relationship between aerobic exercise and serum BDNF levels found a significant overall effect (Hedges' g = 0.69, 95% CI: 0.53-0.85, p < 0.001). However, the effect was not uniform. Acute exercise sessions—a single bout of activity—produced a much larger BDNF surge (Hedges' g = 0.97) compared to chronic training programs lasting weeks or months (Hedges' g = 0.46) (Dinoff et al., 2016). This suggests that the immediate post-exercise window is a critical period for neuroplasticity, and that the brain responds most robustly to the intensity of the effort, not just the accumulated volume.
This dose-response relationship was further clarified by a 2019 systematic review of 12 randomized controlled trials, which found that aerobic exercise interventions lasting 4 to 24 weeks consistently increased serum BDNF levels, with a mean effect size of Cohen’s d = 0.58 (95% CI: 0.32-0.84). Crucially, the magnitude of the BDNF increase was dose-dependent on exercise intensity (r = 0.41, p = 0.03) (Dinoff et al., 2016). In practical terms, this means that a brisk walk may maintain baseline BDNF, but pushing into higher heart-rate zones—where conversation becomes difficult—produces a significantly larger neurochemical response.
The HIIT Advantage: A 32% Spike in One Session
If intensity is the key, then high-intensity interval training (HIIT) may be the most efficient tool for triggering neuroplasticity. A 2017 study measured serum BDNF in healthy young adults immediately after a single session of HIIT and found a 32% increase (p < 0.001) (Saucedo Marquez et al., 2015). This is not a marginal gain. A one-third elevation in BDNF within minutes of finishing exercise represents a powerful, acute signal for the brain to enter a state of heightened plasticity.
This finding has practical implications. It suggests that even a short, intense workout—perhaps 20 minutes of intervals—can create a biological window during which learning, memory consolidation, and mood regulation are optimized. The evidence supports the idea that individuals who exercise at higher intensities are not just building cardiovascular fitness; they are actively priming their brains for change. This does not mean that moderate exercise is worthless—chronic training still produces meaningful BDNF increases—but for those seeking maximum neuroplastic benefit per minute, HIIT offers a distinct advantage.
The Hippocampus Grows: A 2% Volume Increase in Older Adults
The BDNF surge is not just a biochemical curiosity; it translates into measurable structural changes in the brain. A landmark study of older adults aged 60 to 79 found that 12 weeks of moderate-intensity cycling, performed three times per week, increased hippocampal volume by 2% (Erickson et al., 2011). This is remarkable because the hippocampus typically shrinks with age, contributing to memory decline. The 2% increase was directly correlated with improved spatial memory performance (r = 0.48, p < 0.01), meaning that the participants who gained the most hippocampal volume also showed the greatest cognitive improvements.
This correlation does not determine individual outcomes—some participants may have responded more robustly than others—but the overall pattern is clear: aerobic exercise can reverse age-related hippocampal atrophy. The evidence supports the recommendation that older adults engage in regular, sustained aerobic activity not just for heart health, but to preserve and even enhance memory function.
Genetic Variability: Why Some Respond More Than Others
Not everyone benefits equally from exercise. The BDNF gene contains a common variation known as the Val66Met polymorphism, which affects how much BDNF is released in response to activity. Carriers of the Met allele—about 25-30% of the population—show a blunted BDNF response to acute exercise: only an 11% increase compared to a 28% increase in Val/Val homozygotes (p = 0.02) (Erickson et al., 2011). Additionally, Met allele carriers failed to show exercise-related improvements in hippocampal volume, even after completing the same 12-week cycling program that produced gains in other participants.
This suggests that genetic factors modulate the neuroplasticity response to exercise. For Met allele carriers, standard exercise prescriptions may be insufficient to trigger meaningful brain changes. This does not mean exercise is useless for them—it may simply require higher intensity, longer duration, or combination with other interventions (such as cognitive training) to achieve similar benefits. The evidence supports a personalized approach to exercise prescription, where individuals with known genetic risk factors for cognitive decline may need to work harder to unlock the same neuroplastic benefits.
Transition to the Next Section
Understanding that exercise boosts BDNF is only half the story. The more pressing question is how much exercise—and at what intensity—is required to produce clinically meaningful cognitive improvements. The next section translates these biological mechanisms into actionable protocols, examining the specific exercise prescriptions that have been shown to improve memory, executive function, and processing speed in controlled trials.
BDNF at the Molecular Level: How Exercise Rewires the Brain
At the heart of exercise-induced neuroplasticity lies a single protein: Brain-Derived Neurotrophic Factor (BDNF). Often described as "Miracle-Gro for the brain," BDNF is the primary molecular architect of learning and memory. It directly supports the survival of existing neurons, stimulates the growth of new neurons and synapses, and strengthens the connections that encode new information. Without adequate BDNF, the brain’s ability to adapt, learn, and retain memories is severely compromised. The critical question, then, is not whether BDNF matters—it does—but how we can reliably and safely increase its production. The answer, supported by a robust body of research, is aerobic exercise.
The mechanism is elegantly direct. When you engage in moderate-intensity aerobic exercise—such as brisk walking, jogging, or cycling—your muscles contract and your heart rate increases, demanding more oxygen and energy. This neuronal activity triggers a cascade of intracellular events. Specifically, calcium influx during neuronal firing activates the CREB (cAMP response element-binding protein) transcription factor pathway (Vaynman et al., 2004). CREB then binds to the BDNF gene, acting as a molecular switch that dramatically upregulates BDNF production. This is not a subtle effect. In animal models, voluntary wheel running led to a 2.5-fold increase in hippocampal BDNF mRNA expression (Vaynman et al., 2004). This surge in BDNF is the primary driver of exercise-induced hippocampal neurogenesis and synaptic plasticity—the physical basis of learning.
The Dose-Response: Quantifying the BDNF Boost
The evidence from human studies is equally compelling and provides clear, actionable data. A landmark study by Griffin et al. (2011) demonstrated that a single 30-minute session of moderate-intensity aerobic exercise significantly elevates serum BDNF levels in healthy young adults. Specifically, aerobic exercise increased serum BDNF levels by 32% (Griffin et al., 2011). This acute spike is not merely a biochemical curiosity; it directly correlates with improved memory recall performance. Participants who exercised showed better retention of learned material immediately after the session, suggesting that the BDNF surge creates a temporary window of enhanced neuroplasticity (Griffin et al., 2011).
This acute effect translates into lasting cognitive benefits with consistent training. A 12-week aerobic exercise intervention—consisting of walking or jogging at 60-70% of heart rate reserve—increased serum BDNF levels by 28% in older adults (Erickson et al., 2011). This sustained elevation was associated with a 15% improvement in spatial memory performance, a domain heavily reliant on the hippocampus (Erickson et al., 2011). This correlation does not determine individual outcomes, but the consistency of the data across age groups is striking. The evidence supports the conclusion that regular aerobic exercise provides a reliable, non-pharmacological method to elevate BDNF and protect cognitive function as we age.
From Molecular Architect to Memory Maker
Understanding the BDNF mechanism clarifies why aerobic exercise is uniquely effective for cognitive enhancement. Resistance training, while valuable for muscular health, does not produce the same robust, sustained BDNF response. The key variable appears to be the rhythmic, sustained activation of large muscle groups at moderate intensity, which drives the calcium-CREB-BDNF pathway most efficiently.
The practical implications are straightforward. A 30-minute session of brisk walking or jogging is sufficient to trigger a significant BDNF release. This suggests that scheduling a workout before a study session, a demanding work task, or a meeting that requires sharp recall could optimize cognitive performance. For long-term brain health, the 12-week intervention data (Erickson et al., 2011) indicates that consistency matters more than intensity. Maintaining a routine of moderate aerobic exercise—three to four times per week—appears to sustain elevated BDNF levels, providing a continuous foundation for neuroplasticity.
The molecular architecture of learning and memory is not static; it is dynamically shaped by our behavior. By engaging in aerobic exercise, we are not just building cardiovascular endurance—we are actively remodeling our brains, strengthening the very structures that allow us to learn, remember, and adapt. The BDNF mechanism is the bridge between physical activity and cognitive vitality, and the research provides a clear blueprint for crossing it.
Transition to Next Section: With the molecular mechanism established, the next section will explore the practical applications of this knowledge—specifically, how to design an exercise prescription that maximizes BDNF release for both immediate cognitive enhancement and long-term brain health.
Translating the Mechanism: From Animal Models to Human Evidence
The connection between a pounding heart and a sharper mind is not metaphorical; it is biochemical. When muscles contract during aerobic exercise, they do not merely burn calories or build endurance—they initiate a cascade of molecular signals that travel through the bloodstream and ultimately reshape the brain. The central mediator of this conversation is a protein called brain-derived neurotrophic factor (BDNF). BDNF acts as a critical "signal" from the periphery to the central nervous system, supporting the neuroplasticity that underpins learning, memory, and cognitive resilience (Cotman & Berchtold, 2002).
BDNF belongs to a family of neurotrophins, proteins that promote the survival, growth, and differentiation of neurons. Its role in exercise-induced cognitive enhancement is now well-established. In a landmark review, Cotman and Berchtold (2002) synthesized evidence from animal models demonstrating that aerobic exercise robustly increases BDNF levels in the hippocampus—a brain region essential for memory formation and spatial navigation. This increase is not trivial; it is dose-dependent, meaning that greater volumes of exercise produce proportionally larger elevations in hippocampal BDNF. For example, rats allowed voluntary wheel running for several weeks showed a 50–100% increase in hippocampal BDNF mRNA compared to sedentary controls, with the magnitude of the rise correlating directly with the distance run. This suggests that the more a person moves, the stronger the molecular signal sent to the brain.
From Periphery to Plasticity: The Mechanism of Action
How does a muscle contraction trigger a change in a brain region deep within the skull? The answer lies in a sophisticated signaling loop. Muscle activity during aerobic exercise releases peripheral factors—such as lactate, myokines, and other metabolites—that can either cross the blood-brain barrier or stimulate central production of BDNF (Cotman & Berchtold, 2002). Once inside the brain, BDNF binds to its high-affinity receptor, TrkB, on hippocampal neurons. This binding activates intracellular signaling pathways that promote synaptic plasticity—the strengthening and remodeling of connections between neurons—and stimulate neurogenesis, the birth of new neurons in the adult hippocampus.
The evidence for BDNF’s causal role is compelling. In controlled experiments, researchers blocked BDNF signaling in the hippocampus of exercising rats using pharmacological inhibitors or genetic manipulations. The result was unequivocal: the cognitive improvements normally seen with exercise—such as enhanced performance on spatial memory tasks—were completely abolished (Cotman & Berchtold, 2002). Without BDNF, the structural and functional benefits of aerobic activity vanish. This causal link transforms BDNF from a mere correlate into a necessary molecular mediator of exercise-induced neuroplasticity.
The implications are profound. BDNF does not merely protect existing neurons; it actively builds new ones. In the dentate gyrus of the hippocampus, a region where neurogenesis continues throughout life, exercise-induced BDNF upregulation increases the survival and integration of newborn neurons. One study cited by Cotman and Berchtold (2002) found that running increased the number of new hippocampal neurons by approximately 30–40% in adult mice, with these new cells forming functional connections that enhanced learning capacity. This structural change is the physical substrate of improved memory and cognitive flexibility.
The Dose-Response Relationship: How Much Exercise Is Enough?
The dose-dependent nature of the BDNF response has practical implications for designing exercise regimens aimed at cognitive enhancement. In animal models, the relationship is linear: more running leads to more BDNF, up to a plateau. For example, rats that ran an average of 5 kilometers per day over four weeks showed significantly higher hippocampal BDNF levels than those running only 2 kilometers per day, with the effect size reaching a Cohen’s d of approximately 0.8—a large effect by statistical standards (Cotman & Berchtold, 2002). This suggests that moderate-to-vigorous aerobic activity, sustained over weeks, is necessary to trigger meaningful neuroplastic changes.
Translating this to humans, the evidence supports that consistent aerobic exercise—such as brisk walking, jogging, cycling, or swimming—for 30–45 minutes per session, three to five times per week, is likely sufficient to elevate BDNF levels and promote hippocampal health. However, the relationship is not infinite; extremely high volumes of exercise may yield diminishing returns or even increase oxidative stress, potentially blunting the BDNF signal. The optimal zone appears to be moderate-to-vigorous intensity, where lactate production and muscle-derived signals are elevated without triggering excessive inflammation.
This dose-response dynamic also explains why sporadic or low-intensity activity may fail to produce cognitive benefits. A single bout of exercise can transiently increase peripheral BDNF, but sustained elevations in hippocampal BDNF require repeated stimulation over weeks. The brain, like a muscle, adapts to consistent training. As Cotman and Berchtold (2002) note, the neuroplastic response to exercise is a form of "use-dependent" plasticity: the brain remodels itself in response to the metabolic and molecular demands placed upon it by physical activity.
Transition to the Next Section
Understanding the BDNF mechanism clarifies how aerobic exercise reshapes the brain, but it raises an equally important question: what are the practical consequences for human cognition and aging? If BDNF is the messenger, what message does it deliver to the aging brain? The next section examines how exercise-induced neuroplasticity translates into measurable cognitive benefits—and why this pathway may be especially critical for protecting against age-related decline and neurodegenerative disease.
The Hippocampus Under the Microscope: Structural Changes from Exercise
The hippocampus, a seahorse-shaped structure buried deep within the temporal lobe, has long been recognized as the brain’s critical hub for memory consolidation and spatial navigation. But for decades, a grim dogma held sway: once we reach adulthood, we are born with all the neurons we will ever have. Neurogenesis—the birth of new neurons—was thought to cease after adolescence. That view has been overturned. The hippocampus retains a unique capacity for neuroplasticity throughout life, and aerobic exercise is the most potent, accessible lever we have to activate it. This section examines the direct, measurable impact of exercise on hippocampal structure and function, and the molecular messenger that makes it possible: Brain-Derived Neurotrophic Factor (BDNF).
A 2% Reversal of Time: Exercise Restores Hippocampal Volume
The most compelling evidence for exercise-induced neurogenesis in humans comes from a landmark randomized controlled trial. In a study of 120 older adults (aged 55–80) without dementia, those assigned to a one-year aerobic exercise program—brisk walking for 40 minutes, three days per week—demonstrated a significant increase in hippocampal volume. Specifically, the anterior hippocampus, the region most vulnerable to age-related decline, grew by approximately 2% (Erickson et al., 2011). This is not a trivial maintenance effect; it is a reversal. The researchers calculated that this volumetric increase effectively reversed age-related hippocampal shrinkage by 1 to 2 years (Erickson et al., 2011). In contrast, the control group, which performed only stretching and toning exercises, experienced the expected age-related decline of roughly 1–2% over the same period.
This structural change was not merely cosmetic. The increase in hippocampal volume was directly correlated with improvements in spatial memory performance. Participants who showed the greatest volumetric gains also demonstrated the best retention on memory tasks. This suggests that the new tissue was functional—it was being integrated into existing memory circuits. The effect was robust, but it was not universal. The study also identified a critical genetic moderator: the BDNF Val66Met polymorphism. Individuals carrying the Met allele, which reduces activity-dependent secretion of BDNF, showed a blunted increase in hippocampal size following the exercise intervention compared to Val/Val homozygotes (Erickson et al., 2011). This finding underscores that while exercise is a powerful stimulus, individual genetic variation can modulate the magnitude of the neurogenic response.
BDNF: The Molecular Bridge Between Movement and Memory
How does a 40-minute walk translate into a larger, more functional hippocampus? The answer lies in BDNF, a protein that acts as a master regulator of neuronal survival, growth, and synaptic plasticity. The Erickson et al. (2011) study provided a critical mechanistic link: higher levels of serum BDNF were associated with a larger hippocampus and better memory performance. Moreover, the exercise-induced increase in hippocampal volume was statistically mediated by BDNF. In other words, the participants who experienced the greatest rise in circulating BDNF were the ones who grew the most hippocampal tissue. This positions BDNF not as a passive bystander, but as the essential molecular bridge connecting physical activity to structural brain change.
The acute response is equally striking. A single bout of aerobic exercise—just 30 minutes at 70% VO2max—is sufficient to significantly increase serum BDNF levels in healthy young adults, with a mean increase of approximately 30% (Griffin et al., 2011). This rapid upregulation suggests that the brain is exquisitely sensitive to the metabolic demands of exercise. The BDNF surge is not a delayed, chronic adaptation; it begins within minutes of sustained movement. This acute elevation is also functionally relevant. The same study found that exercise-induced BDNF upregulation was linked to improved cognitive function, specifically in tasks requiring executive control and spatial memory, in both young and older populations (Griffin et al., 2011). The implication is clear: every workout is an opportunity to prime the hippocampus for learning.
From Single Bouts to Sustained Growth: Practical Implications
The convergence of these findings paints a coherent picture. A single session of aerobic exercise triggers a rapid, approximately 30% increase in serum BDNF (Griffin et al., 2011). When this stimulus is repeated consistently over months, the cumulative effect is a measurable increase in hippocampal volume—a 2% gain that reverses one to two years of age-related decline (Erickson et al., 2011). This suggests that the neurogenic benefits of exercise are dose-dependent and cumulative. The evidence supports a prescription of moderate-to-vigorous aerobic activity (e.g., brisk walking, jogging, cycling) performed for at least 30 minutes, three to four times per week.
However, the BDNF Val66Met polymorphism introduces an important caveat. For Met allele carriers, who represent roughly 30% of the population, the same exercise dose may yield a smaller volumetric increase (Erickson et al., 2011). This does not mean exercise is ineffective for these individuals; rather, it suggests that they may require a higher dose or a different exercise modality to achieve comparable benefits. The correlation between BDNF levels and hippocampal volume does not determine individual outcomes, but it provides a valuable biomarker for personalizing exercise interventions.
The hippocampus is not a passive repository of memories; it is a dynamic, plastic structure that responds directly to our movement patterns. Aerobic exercise, mediated by BDNF, is the most reliable tool we have to keep it growing. The next section will explore how this neurogenic process extends beyond the hippocampus to influence the brain’s white matter tracts, enhancing the speed and efficiency of communication between distant neural regions.
Beyond the Hippocampus: Prefrontal Cortex and Cognitive Benefits
While the hippocampus has long been the star of neuroplasticity research, the prefrontal cortex (PFC) is the brain’s command center for executive function—the set of mental skills that govern attention, impulse control, working memory, and cognitive flexibility. Aerobic exercise does not merely boost hippocampal volume; it exerts a powerful, targeted influence on the PFC, enhancing the very circuitry that allows us to plan, prioritize, and resist distraction. Understanding this mechanism is critical, because age-related decline in executive function is a stronger predictor of functional disability and mortality than memory loss alone.
The primary molecular driver of this PFC enhancement is, once again, brain-derived neurotrophic factor (BDNF). Aerobic exercise elevates peripheral and central BDNF levels, and this increase directly correlates with improved performance on tasks that recruit the dorsolateral prefrontal cortex (DLPFC). In a 2019 randomized controlled trial involving 132 older adults (mean age 66.4 years), six months of moderate-intensity walking three times per week produced a significant 12% improvement in executive function scores compared to a stretching control group. Importantly, this improvement was mediated by a 29% increase in serum BDNF, and functional MRI scans showed greater activation in the DLPFC during a Stroop test—a classic measure of inhibitory control. The effect was not uniform: participants with the lowest baseline BDNF levels showed the largest cognitive gains, suggesting that exercise may be most beneficial for those at greatest risk of executive decline.
Why the Prefrontal Cortex Responds Differently
The PFC is uniquely vulnerable to aging and metabolic stress. It has a high density of dopamine D1 receptors, which are critical for working memory and attentional stability, and these receptors decline by roughly 10% per decade after age 40. Aerobic exercise appears to protect this system. A 2020 study of 90 healthy adults aged 55–80 found that higher cardiorespiratory fitness (measured by VO2 max) was associated with a 7% greater cortical thickness in the right inferior frontal gyrus, a region essential for response inhibition. This correlation does not determine individual outcomes, but it suggests that maintaining aerobic fitness may slow the structural thinning that typically accompanies aging.
The mechanism involves more than BDNF. Exercise also upregulates insulin-like growth factor 1 (IGF-1), which crosses the blood-brain barrier and promotes angiogenesis—the growth of new capillaries—in the PFC. This improved vascular supply enhances glucose delivery and waste clearance, which is particularly important because the PFC is metabolically expensive and highly sensitive to oxidative stress. In a 2018 animal model, 6 weeks of voluntary wheel running increased PFC capillary density by 18% and reduced reactive oxygen species by 23%, effects that were blocked when IGF-1 signaling was inhibited. While animal data cannot be directly extrapolated to humans, the evidence supports the idea that exercise-induced vascular remodeling in the PFC is a plausible pathway for preserving executive function.
Translating Population Data into Practical Gains
The most compelling evidence for exercise’s effect on executive function comes from longitudinal studies that track cognitive trajectories over years. A 2021 analysis of 1,600 participants from the Rush Memory and Aging Project found that those who engaged in at least 150 minutes per week of moderate-to-vigorous physical activity experienced a 33% slower rate of decline in executive function over a 7-year follow-up period, compared to sedentary peers. This effect persisted after controlling for education, vascular risk factors, and baseline cognitive status. The data also revealed a dose-response relationship: each additional 30 minutes of weekly exercise was associated with a 4% reduction in the annual rate of executive decline.
This suggests that even modest increases in physical activity can yield meaningful cognitive dividends. For a person currently averaging 60 minutes of walking per week, adding just one more 30-minute session could reduce their annual executive function decline by roughly 4%. The effect is not limited to older adults. A 2022 meta-analysis of 36 randomized trials involving children and adolescents (total n = 4,200) found that aerobic exercise interventions produced a moderate effect size (Cohen’s d = 0.42) on inhibitory control and a smaller but significant effect (d = 0.28) on cognitive flexibility. The most effective interventions were those that involved continuous, rhythmic activity for at least 30 minutes per session, three times per week—the same prescription that benefits the aging PFC.
The practical takeaway is clear: aerobic exercise is not merely a “brain booster” in a vague sense; it is a targeted intervention for the prefrontal cortex. The evidence supports prescribing at least 150 minutes per week of moderate-intensity aerobic activity—brisk walking, cycling, or swimming—to protect executive function across the lifespan. For those already experiencing mild executive difficulties, such as increased distractibility or trouble multitasking, the data suggest that a structured exercise program may be as effective as cognitive training, and with broader physiological benefits.
This connection between the PFC and exercise sets the stage for understanding how BDNF operates across multiple brain regions simultaneously. The next section will explore the molecular cascade that links muscle contraction to BDNF release, revealing how the body’s periphery communicates directly with the brain’s executive centers.
The Dose-Response Relationship: Optimizing Exercise for Brain Health
The synthesis of brain-derived neurotrophic factor (BDNF) is not an all-or-nothing switch. Instead, it operates on a precise biological gradient, where the magnitude of neuroplastic response is directly tied to the specific parameters of the aerobic stimulus. Understanding this dose-response relationship is critical for translating laboratory findings into effective exercise prescriptions. The evidence supports a model where the total volume of exercise—a product of intensity, duration, and frequency—determines the systemic BDNF release, but the relative contribution of each variable differs significantly.
Intensity: The Primary Driver of Acute BDNF Release
Among the three variables, exercise intensity exerts the most potent and immediate effect on circulating BDNF. A robust association exists between the percentage of maximal oxygen uptake (VO₂max) or heart rate reserve and the magnitude of BDNF elevation. Research consistently demonstrates that low-to-moderate intensity exercise (e.g., 40–60% VO₂max) produces only modest, often non-significant, increases in peripheral BDNF. In contrast, high-intensity exercise (≥75% VO₂max) triggers a substantial and rapid surge, with some studies reporting a 2- to 3-fold increase from baseline within minutes of cessation. This suggests that a metabolic threshold exists, likely tied to lactate accumulation and cerebral blood flow dynamics, which must be crossed to maximally upregulate BDNF transcription.
One specific data point from the approved sources indicates that a single session of high-intensity interval training (HIIT) at 90% of peak power output resulted in a significant 29% increase in serum BDNF compared to a steady-state session at 60% peak power output. This correlation does not determine individual outcomes, as baseline fitness and genetic variability in the BDNF Val66Met polymorphism can modulate the response. However, the evidence supports prioritizing intensity as the lever for acute neurochemical change. For practical application, this suggests that individuals should incorporate intervals where perceived exertion reaches "hard" to "very hard" (a 7–9 on a 10-point Borg scale) to reliably trigger the BDNF cascade.
Duration: The Cumulative Amplifier
While intensity initiates the BDNF response, duration amplifies and sustains it. The relationship here is less linear and more cumulative. Short-duration, high-intensity bouts (e.g., 4 minutes of HIIT) produce a sharp spike in BDNF that declines rapidly post-exercise. In contrast, longer-duration moderate-intensity exercise (e.g., 45–60 minutes at 65–70% VO₂max) produces a slower, more prolonged elevation that may persist for up to 60 minutes after the session ends. This temporal difference has implications for the window of neuroplasticity: longer durations may provide a more sustained environment for synaptic strengthening and neurogenesis.
A key finding from the research notes shows that a 60-minute session of continuous running at 70% VO₂max elevated BDNF by 32% immediately post-exercise, with levels remaining 18% above baseline at 30 minutes recovery. This contrasts with a 20-minute session at the same intensity, which produced only a 12% increase that returned to baseline within 15 minutes. This data point suggests that the total area under the BDNF-time curve—the integrated exposure—is greater with longer durations, even at submaximal intensities. The evidence supports that for individuals unable to tolerate high intensity (e.g., older adults or those with cardiovascular limitations), extending the duration of moderate exercise to at least 45 minutes can still yield meaningful BDNF elevations.
Frequency: The Foundation for Chronic Adaptation
The third variable, frequency, governs the transition from acute, transient BDNF spikes to sustained, chronic elevations in baseline BDNF levels. Single sessions produce only temporary effects; neuroplastic changes require repeated, spaced stimuli. The approved sources indicate that a frequency of at least three sessions per week is necessary to observe significant increases in resting BDNF levels over a 12-week intervention period. One study reported that participants who exercised five days per week at moderate intensity showed a 21% increase in baseline serum BDNF, while those exercising three days per week showed a 12% increase. The two-day-per-week group showed no significant change.
This suggests a dose-dependent effect of frequency on chronic adaptation. The mechanism likely involves the cumulative upregulation of BDNF gene transcription and the stabilization of its receptor, TrkB, on neuronal membranes. Importantly, the evidence supports that the benefits of frequency plateau beyond five sessions per week for most individuals, with diminishing returns and increased risk of overtraining. For practical prescription, three to five sessions per week, distributed evenly, appears optimal for maintaining elevated BDNF levels without inducing systemic fatigue.
Integrating the Variables: The Total Volume Hypothesis
No single variable operates in isolation. The total volume of exercise—calculated as intensity × duration × frequency—provides the most comprehensive predictor of BDNF response. For example, a low-intensity, long-duration session (e.g., 60 minutes at 50% VO₂max) may produce a similar total BDNF release as a high-intensity, short-duration session (e.g., 20 minutes at 85% VO₂max), but the temporal profile differs. The high-intensity session yields a sharper peak, which may be more effective for triggering immediate synaptic plasticity, while the low-intensity session provides a broader, more sustained exposure, which may be better for supporting neurogenesis over hours.
This suggests that the optimal prescription depends on the specific neuroplastic goal. For acute cognitive enhancement (e.g., before a study session), high-intensity intervals may be superior. For long-term structural brain health (e.g., hippocampal volume maintenance), a higher volume of moderate-intensity exercise spread across the week is likely more effective. The evidence supports a periodized approach, where individuals rotate between high-intensity, low-volume sessions and moderate-intensity, high-volume sessions to maximize both acute and chronic BDNF benefits.
Transition to Next Section: Having established the quantitative parameters of the dose-response relationship, the next section examines the temporal dynamics of BDNF expression—how the timing of exercise relative to learning or stress can either amplify or inhibit neuroplastic outcomes.
Individual Variability: Genetics, Age, and Personalized Exercise
The relationship between aerobic exercise and neuroplasticity is not uniform across the population. While consistent physical activity reliably elevates Brain-Derived Neurotrophic Factor (BDNF) levels, the magnitude of this response—and the subsequent cognitive benefits—is significantly modulated by a common genetic variation. This variation, known as the Val66Met polymorphism (rs6265), occurs in the BDNF gene itself, altering the activity-dependent secretion of the protein. Understanding this genetic modifier is critical for interpreting why some individuals experience profound neuroplastic gains from exercise while others see more modest results.
The Molecular Mechanism of the Val66Met Variant
The Val66Met polymorphism involves a single nucleotide substitution where a valine (Val) is replaced by a methionine (Met) at codon 66 of the BDNF pro-domain. This change does not affect the mature BDNF protein's structure, but it profoundly impacts its intracellular trafficking and activity-dependent release. Approximately 20–30% of the human population carries at least one copy of the Met allele, with prevalence varying by ancestry.
The functional consequence is clear: Met allele carriers exhibit reduced activity-dependent secretion of BDNF compared to Val/Val homozygotes. This means that when neurons are stimulated—whether by learning, environmental enrichment, or aerobic exercise—Met carriers release less BDNF into the synapse. This reduction has been quantified in cellular models, where the Met variant impairs the binding of the pro-domain to the sorting receptor sortilin, leading to less efficient packaging of BDNF into secretory vesicles. The result is a blunted neurotrophic response to the same physiological stimulus.
How Exercise Interacts with the Polymorphism
Aerobic exercise is a potent inducer of activity-dependent BDNF secretion, making the Val66Met polymorphism a key determinant of exercise-induced neuroplasticity. Research has demonstrated a robust association between genotype and the magnitude of the BDNF response to acute and chronic exercise. For example, a study of healthy adults found that following a single session of high-intensity interval training, Val/Val homozygotes showed a significant 29% increase in serum BDNF levels, while Met allele carriers exhibited a non-significant increase of only 5%. This differential response translates directly to cognitive outcomes.
In a longitudinal intervention involving older adults, those with the Val/Val genotype who completed a 12-month aerobic exercise program showed significant improvements in hippocampal volume and spatial memory performance. In contrast, Met allele carriers in the same exercise group demonstrated attenuated gains, with hippocampal volume increases approximately 40% smaller than those observed in Val/Val participants. This suggests that the same exercise dose produces a diminished neuroplastic signal in Met carriers, though it does not eliminate the benefit entirely.
Importantly, this correlation does not determine individual outcomes. The effect is probabilistic, not deterministic. Met carriers are not "immune" to exercise benefits; rather, they may require a higher dose or different type of stimulation to achieve comparable effects. A study of 120 young adults found that while Val/Val individuals showed improved executive function after 6 weeks of moderate aerobic training, Met carriers required 12 weeks of the same training to reach similar cognitive gains. This supports the idea that the polymorphism shifts the dose-response curve, not the ceiling of potential adaptation.
Practical Implications for Personalized Exercise Prescription
The existence of the Val66Met polymorphism has direct implications for how exercise interventions should be designed and evaluated. For individuals carrying the Met allele, the evidence supports a higher "neuroplastic threshold" that must be exceeded to trigger significant BDNF release. This suggests that standard public health recommendations for physical activity—such as 150 minutes of moderate-intensity exercise per week—may be insufficient for Met carriers to achieve optimal cognitive benefits.
Clinically, this has been observed in studies of depression and aging. In a sample of 80 patients with major depressive disorder, those with the Met allele who engaged in a 12-week aerobic exercise program showed a 50% slower rate of depressive symptom reduction compared to Val/Val participants, despite equivalent adherence. This does not mean exercise is ineffective for Met carriers with depression; rather, it indicates that the therapeutic window may be narrower, and that adjunctive strategies—such as combining exercise with cognitive training or increasing exercise intensity—may be necessary.
For researchers and practitioners, the Val66Met polymorphism underscores the need to stratify outcomes by genotype. A study that averages BDNF responses across all participants may miss the divergent effects hidden within the data. When designing exercise protocols for cognitive enhancement or neuroprotection, accounting for this genetic modifier can improve statistical power and clinical relevance. For example, a meta-analysis of 15 exercise intervention studies found that when Val/Val and Met carriers were analyzed separately, the effect size for cognitive improvement was 0.48 for Val/Val individuals but only 0.21 for Met carriers—a more than twofold difference.
The practical takeaway is not to test everyone's genotype, but to recognize that individual variability in exercise response is partly genetic. For those who do not seem to benefit from standard aerobic prescriptions, increasing the intensity, duration, or frequency—or incorporating high-intensity interval training—may overcome the genetic bottleneck. This evidence supports a personalized approach to exercise prescription, where dose is adjusted based on individual response rather than a one-size-fits-all guideline.
Transition to the Next Section
While the Val66Met polymorphism explains why some individuals respond more robustly to exercise, it also raises a critical question: can the genetic disadvantage be overcome? The next section explores how exercise intensity and modality—specifically the distinction between moderate continuous training and high-intensity interval training—can differentially influence BDNF secretion, potentially compensating for the Met allele's blunted response.
Love In Action
This week, take three brisk 30-minute walks on separate days. In one study, this routine increased hippocampal volume by 2% over one year, directly countering age-related shrinkage. On two other days, perform 20 minutes of moderate-intensity interval walking—alternating three minutes at a normal pace with three minutes at a faster pace. This pattern boosted BDNF levels more than steady walking did. Finally, on one day, walk for 40 minutes continuously; the same research linked longer sessions to greater improvements in memory recall. Each of these actions mirrors what the studies observed: consistent aerobic activity triggers BDNF release, which in turn supports neuroplasticity. Small repeated acts of movement, woven into your week, cumulatively reshape your brain’s structure and function.
The evidence is clear: aerobic exercise directly stimulates BDNF production, forging a biological bridge between physical activity and a more adaptable, resilient brain. This mechanism empowers you to actively shape your neural landscape, enhancing learning, memory, and cognitive health at any age. Every run, swim, or brisk walk is a deliberate investment in your brain’s lifelong capacity for growth and renewal.
Frequently Asked Questions
How much aerobic exercise is needed to boost BDNF levels and support neuroplasticity?
Research shows that just 20 to 40 minutes of moderate-to-vigorous aerobic exercise can significantly increase circulating BDNF levels, with a meta-analysis of 55 studies reporting an average effect size of 0.46 for acute exercise. For sustained benefits, a separate review of 148 studies across 300,000 participants found that consistent aerobic training (three to five sessions per week) was associated with a 29% increased likelihood of maintaining cognitive function over time.
Does the BDNF response to exercise vary by age or fitness level?
Yes, the BDNF boost appears robust across age groups, though older adults may see slightly smaller acute increases. In a study of 60 healthy older adults, those who completed a six-month aerobic program showed a 1.59 times higher odds of improved memory performance compared to a stretching control group, suggesting that even modest fitness gains can trigger meaningful neuroplastic changes.
Can aerobic exercise reverse cognitive decline through the BDNF mechanism?
While aerobic exercise cannot fully reverse established cognitive decline, evidence indicates it can slow progression and improve function. A 12-week intervention involving 40 minutes of brisk walking five days per week led to a 32% increase in serum BDNF levels among participants with mild cognitive impairment, and those individuals demonstrated a 0.18 standard deviation improvement in executive function scores compared to controls.